
Con il termine tessuto connettivo si definisce un tipo di tessuto che fornisce supporto strutturale e metabolico agli altri tessuti, per questo motivo è stato proposto il termine di “tessuto di supporto”.
Il tessuto connettivo (e quindi tutte le sue diverse tipologie) deriva dal mesenchima. Le cellule mesenchimali, di origine mesodermica, sono di forma irregolare, generalmente allungate e presentano una sostanza intercellulare amorfa, priva di fibre.
Le cellule mesenchimali sono pluripotenti, hanno cioè la capacità di differenziarsi in ciascuno dei diversi tipi cellulari connettivali: fibroblasti, condrociti, osteociti, mastociti, adipociti, eritrociti (globuli rossi), leucociti (globuli bianchi) e macrofagi, oltre che in fibrocellule muscolari (tessuto muscolare).
Il tessuto connettivo è formato da cellule e da abbondante matrice extracellulare interposta tra di loro. La natura di questa matrice extracellulare determina le caratteristiche dei diversi tipi di tessuto connettivo: si va da una matrice extracellulare fluida (sangue e linfa) in cui possono essere trasportati diversi tipi di cellule connettivali (eritrociti, leucociti, ecc.), ad una matrice solida ma lassa che permette il passaggio di vasi sanguigni e sostanze, fino ad una matrice extracellulare calcificata (tessuto osseo) che permette di creare strutture molto resistenti. Abbiamo quindi una varietà di tipologie di tessuto connettivo che si distinguono sia per il tipo di cellule che le compongono che per la natura della matrice extracellulare presente. Questa matrice extracellulare, detta anche “sostanza intercellulare”, è formata da due tipi di componenti: le fibre (formate da collagene ed altri tipi di proteine) e la sostanza amorfa o fondamentale (formata da proteoglicani). La natura, la morfologia e la consistenza delle fibre e della sostanza amorfa varia notevolmente nelle varie tipologie di connettivo a seconda delle funzioni proprie del tessuto, dell’età dell’individuo e delle caratteristiche particolari della specie presa in esame.
La diversa organizzazione strutturale del tessuto connettivo riflette le sue funzioni.
Quando prevale la funzione trofica, di trasporto di sostanze utili all’organismo e di difesa dalle aggressioni virali o batteriche, il tessuto connettivo deve essere facilmente penetrabile quindi con la struttura leggera, ricca di vasi e dotti linfatici tipica del tessuto connettivo lasso.
Se, invece, la funzione è quella di contenimento, di sostegno o, come nella cute, di una vera e propria barriera, il tessuto connettivo si presenta denso, molto compatto, con fasci di fibre organizzate e orientate in maniera opportuna in modo da assolvere le proprie funzioni, organizzazione tipica del tessuto connettivo denso.
Anche l’organizzazione e la natura delle fibre varia in base alla funzione del tessuto connettivo preso in esame. Al di sotto di tutti gli epiteli, siano essi semplici o stratificati, è presente un connettivo lasso ricco di fibre reticolari che, oltre a formare una base di sostegno per l’epitelio, permette la diffusione di sostanze dai capillari alle cellule epiteliali. Se il tessuto connettivo deve resistere a trazione o a distensione, come nei tendini, è ricco di fibre collagene e di fibre elastiche disposte ordinatamente. Se il connettivo deve essere resistente alla compressione o circondare un grosso organo, la componente principale costituente le fibre connettivali è il collagene.
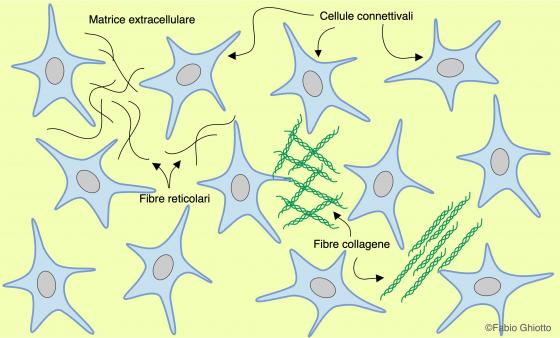
Figura C1. disegno schematico dell’organizzazione del tessuto connettivo.
Fibre collagene
Sono secrete dai fibroblasti e sono presenti nella maggior parte dei tessuti connettivi. Sono formate da una famiglia di proteine secrete come procollagene e poi processate a tropocollagene nell’ambiente extracellulare, tropocollagene che si assembla a formare fibre che variano molto per quantità e che possono apparire come sottili filamenti o come spesse fibre fortemente intrecciate di spessore variabile (fino a 10 µm). La lunghezza delle fibre collagene non è ben determinabile, corrono in tutte le direzioni e, a seconda delle loro funzioni, sono organizzate in fibre parallele, intrecciate o incrociate. Se non sono distese appaiono con un andamento leggermente ondulato.
Le fibre collagene con l’Em-Eo assumono una colorazione rosata, con il Mallory una colorazione azzurro-bluastra. Esistono colorazioni specifiche per le fibre reticolari (collagene di tipo III) ed elastiche. L’intensità di colorazione, in questo tipo di tessuto, è proporzionale al numero e allo spessore delle fibre contenute per cui un tessuto connettivo denso risulterà sempre più colorato di un tessuto connettivo lasso.
| Tipo | Distribuzione | Organizzazione |
|---|---|---|
| I | osso, tendini, dentina, cute | spesse fibre con andamento parallelo o intrecciato, con bande periodiche ben evidenti |
| II | cartilagine ialina e elastica, corpo vitreo | fibre sottili |
| III | tessuto reticolare, connettivo lasso che riveste la muscolatura liscia | fibre sottili che rivestono altri tessuti o che formano trame di molti organi |
| IV | membrane basali | fibre poste al di sotto degli epiteli, sottili e intrecciati al reticolo |
| V | sacco amniotico, corion, guaine muscolari e tendinee | non forma bande |
Tabella C1. Principali tipi di collagene
Fibre elastiche
Le fibre elastiche sono sintetizzate dai fibroblasti, dai condroblasti, dai condrociti e dalle cellule muscolari lisce. Sono formate principalmente dalla proteina elastina. In alcuni organi è molto importate che il tessuto connettivo sia elastico. Le fibre elastiche presenti nel tessuto possono scorrere, allungarsi o deformarsi a causa di sollecitazioni esterne, permettendo quindi l’estensione o la contrazione dell’organo stesso, ma hanno la proprietà di riportare l’organo o il tessuto in cui sono contenute alla situazione originaria. Tessuto connettivo ricco di fibre elastiche si trova, ad esempio, nell’epiglottide, nella vescica urinaria e nella tonaca media delle arterie.
Nelle normali colorazioni topografiche, come l’Em-Eo o l’Azan Mallory, le sottili fibre elastiche non sono facilmente distinguibili dalle altre fibre ma, in fase di riposo e non stirate, possono essere distinte per il loro andamento marcatamente ondulato. Per uno studio accurato sulla presenza di fibre elastiche si deve ricorrere a colorazioni elettive per l’elastina, come la resorcin fucsina di Weigert che, su un fondo praticamente incolore, fa spiccare l’elastina in viola-nero intenso.
Fibre reticolari
Queste fibre formano sia la trama connettivale (stroma) che sostiene il parenchima delle grosse ghiandole, sia esocrine che endocrine, che i sottili reticoli che circondano le cellule nervose, quelle adipose e le fibre muscolari. Sono associate alle lamine basali degli epiteli e formano il tessuto di sostegno degli organi linfatici ed emopoietici.
Anche le fibre reticolari sono difficilmente distinguibili dalle altre fibre del connettivo usando colorazioni convenzionali. Per questo motivo, si sfrutta la capacità delle fibre reticolari, se trattate con soluzioni alcaline di sali d’argento, di legare argento metallico. Vi sono molte colorazioni a base di argento metallico specifiche per le fibre reticolari, una delle più usate è quella di Bielschowsky.
Sostanza fondamentale
Le cellule e le fibre del tessuto connettivo sono immerse in un liquido colloidale e vischioso amorfo denominato sostanza fondamentale o sostanza intercellulare amorfa. La sostanza fondamentale è una rete tridimensionale, più o meno fitta, formata da glicosaminoglicani (GAG), glicoproteine e aggregati di proteoglicani che hanno la capacità di legare acqua e di rendere così la matrice permeabile a sostanze metaboliche e gas che dal sangue passano alle cellule dei tessuti e viceversa. La matrice amorfa e i liquidi tissutali sono, perciò, strettamente associati e svolgono la funzione trofica tipica, ad esempio, del connettivo lasso. La sostanza amorfa adempie a molteplici funzioni: connette le diverse strutture fibrose dando loro un orientamento appropriato; regola la diffusione di sostanze metaboliche; provvede alla difesa dell’organismo ostacolando la diffusione di sostanze nocive e patogeni. La sostanza intercellulare dei tessuti connettivali, con l’età, subisce importanti alterazioni a causa di fenomeni legati alla senescenza.
Durante le preparazioni istologiche questa sostanza si solubilizza ed è quindi impossibile ritrovarla colorata nei preparati. Al criostato, tagliando a circa -30°C e colorando con la PAS reazione o con coloranti basici, si colora molto debolmente. Nella cartilagine e nel tessuto osseo, il ricco contenuto di glicosaminoglicani (GAG) e la consistenza della matrice amorfa permette una buona resistenza ai fissativi e ai solventi chimici, questo permette una buona colorazione con l’Alcian Blu, grazie alla capacità di questo colorante di legare i condroitinsolfati (un tipo di GAG). L’Ematossilina ferrica, come colorazione istologica, e l’Alcian-Pas, come colorazione istochimica, sono le colorazioni che normalmente si usano per dimostrare la sostanza fondamentale. L’Ematossilina ferrica mette in evidenza, non selettivamente, la sostanza intercellulare presente tra le cellule epiteliali o tra le fibre connettivali con una colorazione nera intensa. L’Alcian-Pas è una doppia colorazione istochimica per i glicosaminoglicani: l’Alcian mette in evidenza, con una colorazione blu intensa, quelli acidi, mentre la Pas, con il reattivo di Schiff, mette in evidenza quelli neutri colorandoli in rosso-rosa. La cartilagine ialina matura è colorata molto intensamente dall’Alcian Blu per la presenza di condroitinsolfato ed eparansolfato, entrambi molto acidi.
Il tessuto connettivo si divide in tre categorie principali:
- tessuto connettivo mucoso,
- tessuto connettivo propriamente detto
- tessuti connettivi altamente specializzati
Nota: diversi testi possono riportare una classificazione del tessuto connettivo differente, ad esempio in alcuni testi il tessuto adiposo può essere classificato come tessuto connettivo propriamente detto.